Education for runners and endurance athletes. Learn to train smart, run fast, and be strong.
These articles are free.
Please help keep them alive by buying me a beer:
This article is part of a series:
→ Part 1 — How much fuel is in your body?
→ Part 2 — How do your muscles burn fuel?
→ Part 3 — How long can you go?
→ Part 4 — Carboloading
→ Part 5 — Race day carb availability
→ Part 6 — Putting it into practice
→ Part 1 — How much fuel is in your body?
→ Part 2 — How do your muscles burn fuel?
→ Part 3 — How long can you go?
→ Part 4 — Carboloading
→ Part 5 — Race day carb availability
→ Part 6 — Putting it into practice
Performance nutrition: Part 1 of 6:
How much fuel is available in your body to produce energy during exercise?
Thomas Solomon PhD.
8th May 2021.
I previously went deep into nutritional manipulations for your training. Now it is time to think about how to maximise your performance with nutritional support to keep you speedy on race-day. During this series, I will take you on a journey to help you learn how much fuel your body stores, how your muscles use these fuels during exercise, how to keep energy availability high when you are working hard, and what a fuel plan might look like. To do so, I will even take you back to the 1923 Boston marathon where the need for during-race nutrition began to emerge. So, buckle up and come get your nerd on.
Reading time ~14-mins (2700-words).
Or listen to the Podcast version.
Or listen to the Podcast version.
If you are training to compete, that is to be as fast as you can over a given distance, off the bat, I will be bold and state that “no matter how “fat adapted” you are nor which kind of diet you typically eat, your maximal race-day performance will be achieved with a high carbohydrate availability”. On the contrary, if you choose to line up on race day with a low carbohydrate availability expecting to unleash your fastest performance, you will negate all the hard work you’ve put in to get to that start line.
So, what does that mean? Well, first of all, let’s consider an important question...
 Amino acids, directly from amino acids circulating in blood and from the amino acids that are broken down from functional or structural proteins in various tissues,
Amino acids, directly from amino acids circulating in blood and from the amino acids that are broken down from functional or structural proteins in various tissues,
 Fatty acids, directly from fatty acids circulating in blood and from fatty acids that are broken down from the triglycerides circulating in blood or those stored in adipose tissue, and
Fatty acids, directly from fatty acids circulating in blood and from fatty acids that are broken down from the triglycerides circulating in blood or those stored in adipose tissue, and
 Glucose, directly from glucose circulating in blood and from the glucose that is broken down from stored glycogen in muscle and liver.
Glucose, directly from glucose circulating in blood and from the glucose that is broken down from stored glycogen in muscle and liver.
To conceptualise the relevance of what you have available during endurance exercise, it is useful to think about how much of these fuels are present in your body. So, consider that when the average human wakes up each day their body contains about:
 ~8 kilograms of protein, mostly found in tissues but some protein and amino acids also circulate in the blood. This is equivalent to ~30,000 kcals of energy, enough to fuel an entire day and night of giving it medium-large (running at 80% of your VO2max, i.e. just below your lactate threshold).
~8 kilograms of protein, mostly found in tissues but some protein and amino acids also circulate in the blood. This is equivalent to ~30,000 kcals of energy, enough to fuel an entire day and night of giving it medium-large (running at 80% of your VO2max, i.e. just below your lactate threshold).
Plus, there is about,
~10 kilograms of fat (e.g. in a 65 kg person with 15% body fat), most of which is stored in adipose tissue, with small amounts of triglycerides (~7.5 grams) and fatty acids (~0.01 grams) circulating in blood plus tiny amounts in muscle cells. This is equivalent to ~100’000 kcals of energy, enough to stoke your fire for 3- to 4-days of continuously giving it medium-large (running at 80% VO2max).
And then there is glucose, of which,
~400 grams of glucose is stored in muscle glycogen, equivalent to ~1600 kcals of energy, enough to feed your flame for about ~80-minutes of giving it medium-large.
~100 grams of glucose is stored in liver glycogen, equivalent to ~400 kcals of energy, enough to burn your wick for about ~20-minutes of giving it medium-large.
~4 grams of glucose circulating in the blood, equivalent to ~16 kcals, enough to power Nicolas Cage giving it medium-large (i.e. Gone in 60-seconds, running at 80% VO2max).
Note that other tissues also have glycogen stores but they are very small in comparison to muscle and liver and their relevance during exercise is unknown. For example, astrocytes and neurons in the brain store glycogen but at levels about 100-times lower than in muscle.
Our bodily stores of glucose (glycogen) and fatty acids (triglycerides in adipose tissue) specifically exist to provide these metabolites (fuels) for energy production. Importantly, we do not have a similar protein store that releases amino acids for energy production. Instead, when our body uses amino acids as fuel, it is essentially diverting amino acids away from their essential purpose, which is to build functional or structural proteins. This is why amino acids only contribute around 5 to 10% of total energy production, whereas about 90 to 95% of the energy required to power endurance exercise comes from either glucose or fatty acids — carbohydrates or fats.
When examining the relative amounts of fuel substrates available in the body during exercise, there are three things to note… Firstly, the average human has enough protein in the body to keep burning for days. Secondly, the average human stores a huge amount of fat — about 10,000 grams or 100,000 kcals — even the fat stored in a super lean athlete will not run out. Thirdly, the average human stores very little carbohydrate — about 500 grams or 2000 kcals — which can be “burned” through fast.
So, you can paint a tasty picture of the fuels available to you during endurance exercise:
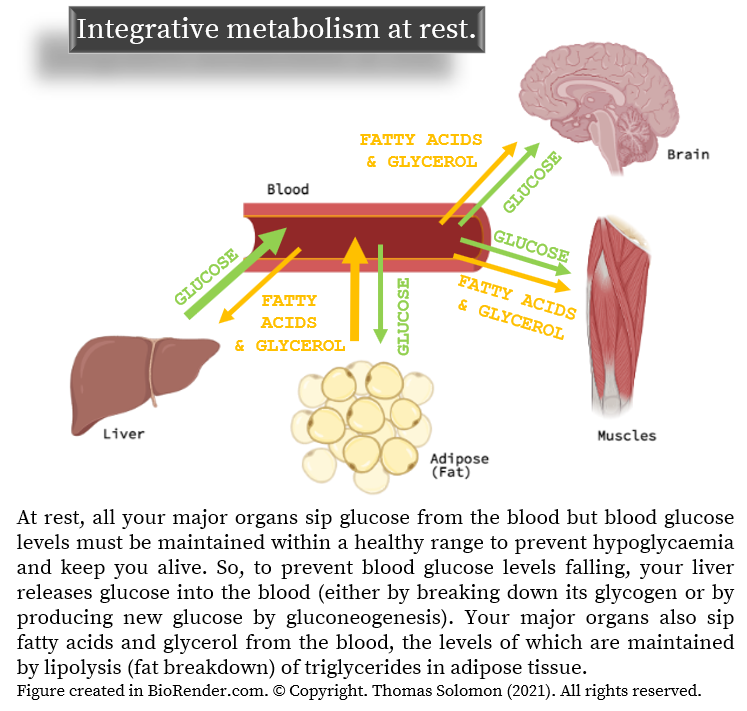
Your blood contains about 4 grams of glucose circulating around the body. In healthy people, the blood glucose concentration (the amount of glucose per litre of blood) is very tightly regulated at approximately 1 gram per litre or 5 mmol/L. (NOTE: these are not precise amounts, just estimates. A healthy fasting blood glucose concentration falls in the range of 4.0 to 5.6 mmol/L, equivalent to 0.7 to 1.0 g/L or 72 to 100 mg/dL.) Of all the many “things” dissolved in your blood, glucose is relatively abundant and therefore greatly contributes to something called osmolarity, which you can think of as a “liquid pressure” that blood exerts allowing all of the “things” to leave the blood and enter tissues, and vice versa, at appropriate rates. When glucose levels change, even slightly, so too does this “liquid pressure”. So, the body must quickly restore homeostasis to prevent bodily functions from going haywire and to keep you healthy. For example, eating a meal makes the blood glucose concentration rise (perhaps to 15 g/L, or 8 mM) but, in healthy people, it recovers to ~1 g/L (5 mM) within a couple of hours. This transient rise and fall in glucose is fine and has no dire consequences. Plus, a healthy body has homeostatic mechanisms that prevent prolonged hypoglycaemia (low glucose, less than 4 mM) or hyperglycaemia (high glucose, more than 8 mM), the direct consequences of which include oxidative stress, inflammation, and electrolyte disturbances that have direct detrimental effects on your major organs, including your heart, brain, liver, and kidneys, and also your muscles.
Although the level of glucose in your blood is tightly regulated, all of the metabolic processes in your body constantly need energy (ATP) and the blood delivers fuels that produce the energy to the tissues that need it. This is true even when you rest or sleep because your brain, muscles, kidneys, and red blood cells are particularly hungry glucose-guzzlers and, consequently, glucose is constantly leaving the blood to be used as a fuel to keep your neurons firing and other organs functioning. In fact, at rest, about 8 to 10 grams of glucose leave the blood every hour!
Perhaps you are wondering, “if glucose keeps leaving the blood, surely it must also have to enter the blood from somewhere for blood glucose levels to stay within the healthy range?”. Hell yeah, it does!
During exercise, energy demands dramatically increase because of the many muscle contractions taking place. Your muscles remove lots of glucose from the blood and the amount of glucose leaving the blood every hour jumps from 8 to 10 grams/hour at rest to ~60 to 90-grams per hour during exercise — a 6 to 10-fold increase in the amount of glucose leaving the blood every hour. Your liver must now work like a horse to release sufficient glucose to maintain blood levels at ~1 g/L (5 mM). And, it does a phenomenal job — preventing blood glucose dropping even when exercising muscles are gobbling it up like a hoover. Well, at least for a while. More on that later...
Just like your muscles, your liver also adapts to training by increasing its capacity to metabolise fat to produce the energy it needs to keep supplying the muscle with glucose — amazing! Without a liver, the metabolic demands of contracting muscle would quickly deplete blood glucose (causing hypoglycemia, uh oh), the brain would fail, and your running journey will end... Forever... But you do have a liver, so keep it healthy. To go deeper on the role of the liver in exercise, I can recommend reading the excellent 2016 review by Javier Gonzalez and colleagues.
Now you have a small (and very simplified) insight into the beautiful world of integrative physiology — your adipose tissue supplies fat to the liver so it can maintain blood glucose levels while your brain, muscles, and other tissues guzzle away at the sugary goodness, all the while connected by blood vessels and a pumping heart... and I haven’t even mentioned the role your kidneys, lungs, adrenal glands, and pancreas all play in that process (and nor will I be delving into such things here).
Phew.
So, now you are armed with knowledge of how much fuel is available to you in your body — a bucket load of fatty acids and pint-glass of glucose — but I also hope you are starting to appreciate how your organ systems fit together.
During this series on performance nutrition, I will be delving back in time and taking you on a journey through the ages to help you understand how these pools of fuel are used during exercise. But through this time continuum, there have been important developments in methodology that allow us to peer inside the body to measure metabolite flux (the rate at which a specific fuel, like glucose, goes into and out of a tissue) and metabolite oxidation (the rate at which specific fuels, like glucose, are “burned” to produce chemical energy, ATP). For this reason, it is important to ask yourself:
In the early 1900s, the Nobel prize winner August Krogh pioneered the use of indirect calorimetry to assess whole-body oxidation; that is, the rates of oxygen consumption and carbon dioxide production, measured in your expired breath, at rest and during exercise. After Krogh’s development, many studies examined whole body metabolism coupled with metabolite snapshots in the blood. But something a little more innovative was needed to advance our understanding.
With the exception of some silly wars, the 1960s were pretty groovy baby. We made some pretty cool advances: voting rights, gender rights, one chap even left our glorious third rock from the sun and took a stroll on the moon. But, in the 60s, there were many other non-televised giant leaps for mankind that were far cheaper than sending a rocket into space and have had a far greater impact on our current understanding of metabolic health and disease.
Firstly, arterio-venous (AV) balance methods were developed to study metabolic flux across tissues, i.e. the rate of transport into or out of tissues (like the muscle or liver) from the blood. Because of the technical expertise and high-risk, arterial catheters are typically reserved for clinical diagnostics but AV balance is used in metabolic studies. In the 1960s, tissue biopsies also began to be used to examine tissue-specific levels of metabolites and molecules. For example, Eric Hultman and Jonas Bergström pioneered the use of liver and muscle biopsies in exercise science to measure glucose stores (glycogen) and fat stores. But due to their high-risk, liver biopsies in exercise studies are exceptionally rare, whereas muscle and fat biopsies are commonplace. AV-balance methods and biopsies were (and remain) useful for understanding metabolite movement and metabolite tissue levels, but something more was needed.
Between the 1960s and 2000s, radioisotopes of carbon-14 and hydrogen-3 were commonly used to assess tissue flux and metabolite oxidation without the need for highly invasive arterial catheters. They are less common nowadays because of radiation exposure. But isotope methods are very useful because carbon dioxide (CO2) is a product of metabolism and so, injecting or ingesting a metabolite (like glucose) that carries a “heavy” carbon atom (14C) can be traced through the body and measured as a heavy 14CO2 in expired breath, thus telling us exactly how much of the injected or ingested glucose was oxidized (“burned”).
Pretty clever!
Consequently, non-radioactive stable isotopes of carbon-13 and hydrogen-2 have replaced the need for radioisotopes and are now widespread and have been used since the 1990s to study metabolite flux and oxidation rates. But, it doesn’t stop there — during the 1980s the use of nuclear magnetic resonance spectroscopy in metabolic studies became increasingly popular. Folks like Gerald Shulman have pioneered its use to detect the resonance of hydrogen, phosphorus, and carbon atoms in the presence of a strong magnetic field (like a MRI scan) in muscle and liver glycogen and triglycerides at rest and during exercise, without the need for isotopes or biopsies.
Nerdy.
Now you know how scientists peek inside the living body and measure metabolite flux and oxidation. You also know that you have a lot of stored fuel that is available to “burn” to produce energy during exercise: a bucket load of fatty acids and a pint-glass of glucose.
With these knowledge nuggets, you have the appropriate brain food to tackle a burning question: How does your body use your available “fuels” during exercise?.
Phenomenal question. But, you will have to wait for the next part of this series.
Until then, keep training smart…

So, what does that mean? Well, first of all, let’s consider an important question...
What type of fuel does your body have available for endurance exercise?
As you probably already know, to produce ATP (energy) during endurance exercise, the human body uses:
Amino acids, directly from amino acids circulating in blood and from the amino acids that are broken down from functional or structural proteins in various tissues,
Fatty acids, directly from fatty acids circulating in blood and from fatty acids that are broken down from the triglycerides circulating in blood or those stored in adipose tissue, and
Glucose, directly from glucose circulating in blood and from the glucose that is broken down from stored glycogen in muscle and liver.
Plus, there is about,
And then there is glucose, of which,
Our bodily stores of glucose (glycogen) and fatty acids (triglycerides in adipose tissue) specifically exist to provide these metabolites (fuels) for energy production. Importantly, we do not have a similar protein store that releases amino acids for energy production. Instead, when our body uses amino acids as fuel, it is essentially diverting amino acids away from their essential purpose, which is to build functional or structural proteins. This is why amino acids only contribute around 5 to 10% of total energy production, whereas about 90 to 95% of the energy required to power endurance exercise comes from either glucose or fatty acids — carbohydrates or fats.
When examining the relative amounts of fuel substrates available in the body during exercise, there are three things to note… Firstly, the average human has enough protein in the body to keep burning for days. Secondly, the average human stores a huge amount of fat — about 10,000 grams or 100,000 kcals — even the fat stored in a super lean athlete will not run out. Thirdly, the average human stores very little carbohydrate — about 500 grams or 2000 kcals — which can be “burned” through fast.
So, you can paint a tasty picture of the fuels available to you during endurance exercise:
A bucket load of amino acids,
a bucket load of fatty acids,
and a pint-glass of glucose.
But, because amino acids are not a primary fuel and contribute less than 5 to 10% of total energy even in very long-duration events, in the context of exercise metabolism, we can disregard them. Therefore, we can simplify our sticky picture to:
a bucket load of fatty acids,
and a pint-glass of glucose.
A bucket load of fatty acids
and a pint-glass of glucose.
(Note: some studies indicate that protein breakdown for energy production might become more relevant during extreme ultra-duration events. But, even under such conditions, protein’s contribution is still less than 10% of total energy production and we do not have a clear understanding of if or how this affects performance. I will return to the potential relevance of protein and amino acids in the context of ultra-distance races in a future post.)
and a pint-glass of glucose.
The importance of blood glucose concentrations.
Your fat stores will not run out during exercise but your carbohydrate stores can. I will come back to the relative contributions of the two but there is something else very important to learn to help this whole topic make sense...Your blood contains about 4 grams of glucose circulating around the body. In healthy people, the blood glucose concentration (the amount of glucose per litre of blood) is very tightly regulated at approximately 1 gram per litre or 5 mmol/L. (NOTE: these are not precise amounts, just estimates. A healthy fasting blood glucose concentration falls in the range of 4.0 to 5.6 mmol/L, equivalent to 0.7 to 1.0 g/L or 72 to 100 mg/dL.) Of all the many “things” dissolved in your blood, glucose is relatively abundant and therefore greatly contributes to something called osmolarity, which you can think of as a “liquid pressure” that blood exerts allowing all of the “things” to leave the blood and enter tissues, and vice versa, at appropriate rates. When glucose levels change, even slightly, so too does this “liquid pressure”. So, the body must quickly restore homeostasis to prevent bodily functions from going haywire and to keep you healthy. For example, eating a meal makes the blood glucose concentration rise (perhaps to 15 g/L, or 8 mM) but, in healthy people, it recovers to ~1 g/L (5 mM) within a couple of hours. This transient rise and fall in glucose is fine and has no dire consequences. Plus, a healthy body has homeostatic mechanisms that prevent prolonged hypoglycaemia (low glucose, less than 4 mM) or hyperglycaemia (high glucose, more than 8 mM), the direct consequences of which include oxidative stress, inflammation, and electrolyte disturbances that have direct detrimental effects on your major organs, including your heart, brain, liver, and kidneys, and also your muscles.
Although the level of glucose in your blood is tightly regulated, all of the metabolic processes in your body constantly need energy (ATP) and the blood delivers fuels that produce the energy to the tissues that need it. This is true even when you rest or sleep because your brain, muscles, kidneys, and red blood cells are particularly hungry glucose-guzzlers and, consequently, glucose is constantly leaving the blood to be used as a fuel to keep your neurons firing and other organs functioning. In fact, at rest, about 8 to 10 grams of glucose leave the blood every hour!
Perhaps you are wondering, “if glucose keeps leaving the blood, surely it must also have to enter the blood from somewhere for blood glucose levels to stay within the healthy range?”. Hell yeah, it does!
Your liver is awesome.
To prevent blood glucose levels from dropping too low — to prevent hypoglycaemia — when your tissues are feasting on the bloody but sugary goodness, the liver releases glucose into the blood. It does so by either releasing glucose from its own (small) store of glycogen (glycogenolysis) or by producing new glucose (gluconeogenesis) from other metabolites, like glycerol, pyruvate, lactate, and amino acids. These are energy costly processes that the liver cells fuel by producing energy from circulating fatty acids, which are primarily released into the blood by adipose tissue (fat tissue).Note that although muscles have a glycogen store, they also lack the enzyme that allows glucose to be released into the blood and so the glucose broken down from muscle glycogen is entirely used to produce energy within the muscle — glucose from muscle glycogen is never used to help maintain blood glucose concentrations..
During exercise, energy demands dramatically increase because of the many muscle contractions taking place. Your muscles remove lots of glucose from the blood and the amount of glucose leaving the blood every hour jumps from 8 to 10 grams/hour at rest to ~60 to 90-grams per hour during exercise — a 6 to 10-fold increase in the amount of glucose leaving the blood every hour. Your liver must now work like a horse to release sufficient glucose to maintain blood levels at ~1 g/L (5 mM). And, it does a phenomenal job — preventing blood glucose dropping even when exercising muscles are gobbling it up like a hoover. Well, at least for a while. More on that later...
Just like your muscles, your liver also adapts to training by increasing its capacity to metabolise fat to produce the energy it needs to keep supplying the muscle with glucose — amazing! Without a liver, the metabolic demands of contracting muscle would quickly deplete blood glucose (causing hypoglycemia, uh oh), the brain would fail, and your running journey will end... Forever... But you do have a liver, so keep it healthy. To go deeper on the role of the liver in exercise, I can recommend reading the excellent 2016 review by Javier Gonzalez and colleagues.
Now you have a small (and very simplified) insight into the beautiful world of integrative physiology — your adipose tissue supplies fat to the liver so it can maintain blood glucose levels while your brain, muscles, and other tissues guzzle away at the sugary goodness, all the while connected by blood vessels and a pumping heart... and I haven’t even mentioned the role your kidneys, lungs, adrenal glands, and pancreas all play in that process (and nor will I be delving into such things here).
Phew.
×
![]()
Note: To read more about the liver’s role in exercise metabolism, I can recommend an excellent 2015 review published by Prof David Wasserman and his colleagues at Vanderbilt.
So, now you are armed with knowledge of how much fuel is available to you in your body — a bucket load of fatty acids and pint-glass of glucose — but I also hope you are starting to appreciate how your organ systems fit together.
During this series on performance nutrition, I will be delving back in time and taking you on a journey through the ages to help you understand how these pools of fuel are used during exercise. But through this time continuum, there have been important developments in methodology that allow us to peer inside the body to measure metabolite flux (the rate at which a specific fuel, like glucose, goes into and out of a tissue) and metabolite oxidation (the rate at which specific fuels, like glucose, are “burned” to produce chemical energy, ATP). For this reason, it is important to ask yourself:
How do we measure fuel use during exercise?
Measuring the concentration of a metabolite, say glucose, in the blood at a single time point only provides us with a “Kodak-moment”, aka a snapshot of blood glucose levels at that time. This can be useful for clinical diagnostics but a snapshot tells us nothing about whether the levels are stable, or rising or falling. Neither does a snapshot tell us anything about where glucose has come from or is going to (metabolite flux), nor does it tell us how much glucose is being “burned” (metabolite oxidation) to produce energy (ATP).In the early 1900s, the Nobel prize winner August Krogh pioneered the use of indirect calorimetry to assess whole-body oxidation; that is, the rates of oxygen consumption and carbon dioxide production, measured in your expired breath, at rest and during exercise. After Krogh’s development, many studies examined whole body metabolism coupled with metabolite snapshots in the blood. But something a little more innovative was needed to advance our understanding.
With the exception of some silly wars, the 1960s were pretty groovy baby. We made some pretty cool advances: voting rights, gender rights, one chap even left our glorious third rock from the sun and took a stroll on the moon. But, in the 60s, there were many other non-televised giant leaps for mankind that were far cheaper than sending a rocket into space and have had a far greater impact on our current understanding of metabolic health and disease.
Firstly, arterio-venous (AV) balance methods were developed to study metabolic flux across tissues, i.e. the rate of transport into or out of tissues (like the muscle or liver) from the blood. Because of the technical expertise and high-risk, arterial catheters are typically reserved for clinical diagnostics but AV balance is used in metabolic studies. In the 1960s, tissue biopsies also began to be used to examine tissue-specific levels of metabolites and molecules. For example, Eric Hultman and Jonas Bergström pioneered the use of liver and muscle biopsies in exercise science to measure glucose stores (glycogen) and fat stores. But due to their high-risk, liver biopsies in exercise studies are exceptionally rare, whereas muscle and fat biopsies are commonplace. AV-balance methods and biopsies were (and remain) useful for understanding metabolite movement and metabolite tissue levels, but something more was needed.
Between the 1960s and 2000s, radioisotopes of carbon-14 and hydrogen-3 were commonly used to assess tissue flux and metabolite oxidation without the need for highly invasive arterial catheters. They are less common nowadays because of radiation exposure. But isotope methods are very useful because carbon dioxide (CO2) is a product of metabolism and so, injecting or ingesting a metabolite (like glucose) that carries a “heavy” carbon atom (14C) can be traced through the body and measured as a heavy 14CO2 in expired breath, thus telling us exactly how much of the injected or ingested glucose was oxidized (“burned”).
Pretty clever!
Consequently, non-radioactive stable isotopes of carbon-13 and hydrogen-2 have replaced the need for radioisotopes and are now widespread and have been used since the 1990s to study metabolite flux and oxidation rates. But, it doesn’t stop there — during the 1980s the use of nuclear magnetic resonance spectroscopy in metabolic studies became increasingly popular. Folks like Gerald Shulman have pioneered its use to detect the resonance of hydrogen, phosphorus, and carbon atoms in the presence of a strong magnetic field (like a MRI scan) in muscle and liver glycogen and triglycerides at rest and during exercise, without the need for isotopes or biopsies.
Nerdy.
What can you put in your performance nutrition toolbox?
Today, researchers commonly combine indirect calorimetry with AV-balance, tissue biopsies, and stable isotopes or use magnetic resonance spectroscopy to generate highly-detailed models of human metabolism. I have been very fortunate to use all of these methods in my work.Now you know how scientists peek inside the living body and measure metabolite flux and oxidation. You also know that you have a lot of stored fuel that is available to “burn” to produce energy during exercise: a bucket load of fatty acids and a pint-glass of glucose.
With these knowledge nuggets, you have the appropriate brain food to tackle a burning question: How does your body use your available “fuels” during exercise?.
Phenomenal question. But, you will have to wait for the next part of this series.
Until then, keep training smart…
×
![]()
Disclaimer: I occasionally mention brands and products but it is important to know that I am not affiliated with, sponsored by, an ambassador for, or receiving advertisement royalties from any brands. I have conducted biomedical research for which I have received research money from publicly-funded national research councils and medical charities, and also from private companies, including Novo Nordisk Foundation, AstraZeneca, Amylin, A.P. Møller Foundation, and Augustinus Foundation. I’ve also consulted for Boost Treadmills and Gu Energy on their research and innovation grant applications and I’ve provided research and science writing services for Examine — some of my articles contain links to information provided by Examine but I do not receive any royalties or bonuses from those links. These companies had no control over the research design, data analysis, or publication outcomes of my work. Any recommendations I make are, and always will be, based on my own views and opinions shaped by the evidence available. My recommendations have never and will never be influenced by affiliations, sponsorships, advertisement royalties, etc. The information I provide is not medical advice. Before making any changes to your habits of daily living based on any information I provide, always ensure it is safe for you to do so and consult your doctor if you are unsure.
If you find value in this free content, please help keep it alive and buy me a beer:
Share this post on your social media:
Want free exercise science education delivered to your inbox? Join the 100s of other athletes, coaches, students, scientists, & clinicians and sign up here:

About the author:
I am Thomas Solomon and I'm passionate about relaying accurate and clear scientific information to the masses to help folks meet their fitness and performance goals. I hold a BSc in Biochemistry and a PhD in Exercise Science and am an ACSM-certified Exercise Physiologist and Personal Trainer, a VDOT-certified Distance running coach, and a Registered Nutritionist. Since 2002, I have conducted biomedical research in exercise and nutrition and have taught and led university courses in exercise physiology, nutrition, biochemistry, and molecular medicine. My work is published in over 80 peer-reviewed medical journal publications and I have delivered more than 50 conference presentations & invited talks at universities and medical societies. I have coached and provided training plans for truck-loads of athletes, have competed at a high level in running, cycling, and obstacle course racing, and continue to run, ride, ski, hike, lift, and climb as much as my ageing body will allow. To stay on top of scientific developments, I consult for scientists, participate in journal clubs, peer-review papers for medical journals, and I invest every Friday in reading what new delights have spawned onto PubMed. In my spare time, I hunt for phenomenal mountain views to capture through the lens, boulder problems to solve, and for new craft beers to drink with the goal of sending my gustatory system into a hullabaloo.
Copyright © Thomas Solomon. All rights reserved.
I am Thomas Solomon and I'm passionate about relaying accurate and clear scientific information to the masses to help folks meet their fitness and performance goals. I hold a BSc in Biochemistry and a PhD in Exercise Science and am an ACSM-certified Exercise Physiologist and Personal Trainer, a VDOT-certified Distance running coach, and a Registered Nutritionist. Since 2002, I have conducted biomedical research in exercise and nutrition and have taught and led university courses in exercise physiology, nutrition, biochemistry, and molecular medicine. My work is published in over 80 peer-reviewed medical journal publications and I have delivered more than 50 conference presentations & invited talks at universities and medical societies. I have coached and provided training plans for truck-loads of athletes, have competed at a high level in running, cycling, and obstacle course racing, and continue to run, ride, ski, hike, lift, and climb as much as my ageing body will allow. To stay on top of scientific developments, I consult for scientists, participate in journal clubs, peer-review papers for medical journals, and I invest every Friday in reading what new delights have spawned onto PubMed. In my spare time, I hunt for phenomenal mountain views to capture through the lens, boulder problems to solve, and for new craft beers to drink with the goal of sending my gustatory system into a hullabaloo.
Copyright © Thomas Solomon. All rights reserved.